

Inbreeding is heritable in ...
Human societies generally consider inbreeding, or incestuous mating with a close relative, taboo. Many people also know that inbreeding is often bad for resulting kids, and hence best avoided. However, many wild animals and plants commonly inbreed causing scientists to wonder why individuals don't always avoid incestuous matings?
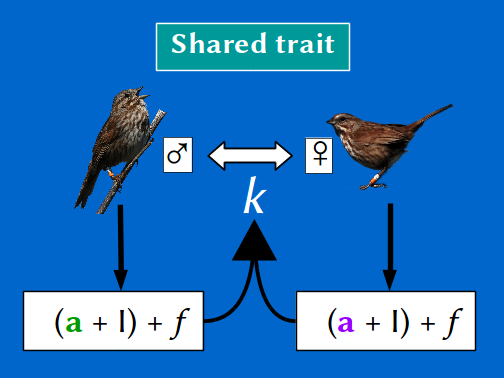
In collaboration with Jane Reid, we used 40 years of data collected from a wild population of song sparrows (Melospiza melodia) inhabiting Mandarte island (British Columbia, Canada) to calculate the degree to which inbreeding was due to individuals' genes versus environment. The song sparrow population provides a unique opportunity to undertake such analyses, because the relatedness among all sparrows hatched in the population since 1975 is known with exceptionally high accuracy.
Biparental inbreeding is ...
A mixed model analysis, including both indirect genetic effects and genetic group effects, was used to model the degree of inbreeding through social pairing as an emergent trait of each pair. We showed that the degree of biparental inbreeing is heritable, implying that the population mean could actually evolve and hence become more or less over time. We also found a high between-sex additive genetic correlation. Consequently, differences in sex-specific selection could lead to evolutionary sexual conflict over the degree of biparental inbreeding (Wolak & Reid 2016).
The results provide new insights into what drives mating between relatives and what the consequences might be for small populations where inbreeding is likely to occur. The findings are a key first step in determining how organisms mate in nature which is essential to knowing how wild populations will adapt to their environment and evolve over time.

Sexual dimorphisms are ...
Phenotypic differences between females and males, or sexual dimorphisms, are widely observed in organisms with separate sexes. Since the sexes share the majority of their genomes, evolutionary biologists are faced with a problem. How do we explain the large differences between the sexes, when mothers pass on female-favorable alleles to their sons and fathers do the same to their daughters?

Work I conducted as ...
Given the many observed instances of the sexes responding to selection in opposite directions, this implies the existence of sex-specific genetic variation. Sex-specific genetic variation facilitates the evolution of sexual dimorphisms. Thus, a major focus of research regarding the evolution of sexual dimorphism is to determine what mechanisms create sex-specific genetic variation within a population and how might these mechanisms differ among populations.

Using line crosses among ...
Current theory suggests sex-linkage of sex-specific genetic effects as a mechanism underlying the genetic architecture of sexually dimorphic traits. Alternatively, it is hypothesized that sex-specific non-additive effects may be a separate mechanism. I have been working in collaboration with Daphne Fairbairn to test these two hypotheses using populations of the water strider (Aquarius remigis).

Despite >70 generations of artificial selection ...
How does selection shape the underlying quantitative genetic architecture and therefore alter future responses to selection? I collaborate with Vincent Careau, Pat Carter, and Ted Garland, Jr. to study the changes in additive genetic (co)variances occurring during long-term artificial selection on behavioral traits in house mice. Experimental evolution is a particularly advantageous tool with which to study change in additive genetic (co)variances, because it enables us to introduce replication and controls in the experimental design. In this experiment, we have 4 selected and 4 control lines of mice. As with any dataset, this replication gives us the ability to make fairly strong inferences.
We have asked how changes in additive genetic variances and covariances might explain apparent limits to univariate and multivariate evolutionary responses to artificial selection on voluntary wheel running. To accomplish this, we have estimated the changes in additive genetic (co)variances that can be attributed to selection, drift, and/or linkage disequilibrium. Future work will investigate how within-individual variance may be heritable and play a role in responses to artificial selection.